A short introduction to Epigenetics
Epigenetics is a normal gene regulatory phenomenon involved in human development and cellular differentiation. Indeed, each cell type in the body is genetically identical but needs to differentiate phenotypically to sustain a normally functioning human body: neurons, skin, etc. However, there is evidence that several external factors can impact gene function epigenetically, both positively and negatively, including aging, environmental exposure, diseases like cancer, diet, and even exercise.
The field of epigenetics is relatively new and needs more research to understand if epigenetic changes actually cause disease or if they are simply correlational. Likely a bit of both. However, several companies, including publicly traded companies, are looking at creating both epigenetically-based diagnostics and therapeutics. Many companies, including startups focused on the male-fertility space are developing diagnostics that will better elucidate problems in male infertility, mental health, and cancer.
Learn more about how epigenetics impacts you
The Early Origins of Epigenetics
Although mechanistically incorrect, Charles Darwin first conceptually described a possible biological process for epigenetic inheritance in 1868 through his theory of Pangenesis. Gemmules, Darwin suggested, were imagined particles of inheritance that were shed from organs of the body and traveled through the bloodstream to the reproductive organs where they would accumulate in the gametes. There, they would integrate with the germ cells to impact future generations; a sort of short-term evolutionary change. Thus, gemmules provided a possible mechanism for the inheritance of acquired characteristics first proposed in 1809 by Jean-Baptiste Lamarck.
Until the second half of the 20th century, human developmental biology and genetics were treated as separate scientific disciplines. Conrad Waddington is given credit for coining the term, epigenetics, in 1942 that brought these two fields together. The term was derived from the Greek word “epigenesis” which originally described the effects of genetic processes on human development. It was several decades later when 5-methyl cytosine was suggested to have a role in controlling gene activity, and that DNA methylation of cytosine bases was heritable. These initial findings are now supported by a significant body of research that show DNA methylation is involved with silencing of gene activity in multiple biological systems. Additionally, other very important epigenetic mechanisms of gene regulation have been discovered including histone modifications and regulatory RNAs.
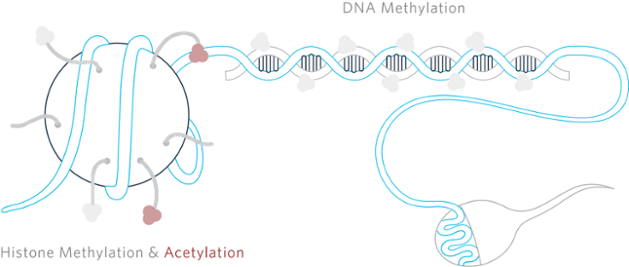
Epigenetic Mechanisms
Individual cells encode their epigenetic information through modifications to either the DNA itself or histones, the proteins that DNA coil around. One other epigenetic mechanism that has recently been discovered is regulating micro RNA, which won’t be discussed here.
DNA Methylation
Currently, DNA methylation is one of the most well-understood epigenetic modifications that dates back to studies conducted by Griffith and Mahler in 1969, which suggested that this methylation of DNA may be important for long-term memory. DNA methylation is a common regulatory mark found in many different cell types on the 5-carbon of cytosine residues (5-mC) at cytosine–phosphate–guanine dinucleotides (CpGs) in DNA. DNA methylation is essential in genomic imprinting, gene expression regulation, X chromosomal inactivation, and embryonic development. This epigenetic mark can activate or repress gene transcription at specific sites based on the methylation levels at promoter regions.
Hypermethylation at promoters blocks access of transcriptional machinery and thus inhibits gene expression. Conversely, hypomethylation facilitates gene activation as a result of increased accessibility of DNA by polymerase. The regulation of DNA methylation is essential to normal cell function in somatic cells, gametes, and the embryo. The DNA methyltransferase (DNMT) family of proteins helps to facilitate de novo methylation and methylation maintenance. The enzymes directly responsible for de novo methylation include DNMT3A, DNMT3B, and DNMT3L. Both DNMT3A and DNMT3B contain catalytic domains, which allow them to directly lay down new methylation marks. DNMT3L is essential in directing the proper placement of marks by working in concert with DNMT3A and DNMT3B. Once methylation marks have been established, DNMT1 maintains those marks through cell division.
Histone Modification
Histones can be modified through a number of post-translational mechanisms including lysine and arginine methylation, serene and threonine phosphorylation, lysine acetylation, and lysine ubiquitination and sumoylation. These modifications occur primarily within the histone amino-terminal tails protruding from the surface of the nucleosome as well as on the globular core region. These different modifications to the histones are thought to impact chromosome function through a couple of different mechanisms. The first mechanism alters the electrostatic charge of the histone and causes a structural change within the histone or their binding to the DNA. The second mechanism suggests modifications to binding sites for protein recognition modules that recognize acetylated or methylated lysines. Like DNA methylation, these posttranslational modifications of histones can create an epigenetic mechanism for the regulation of a variety of normal and disease-related processes.
Impact of Epigenetics in Reproductive Health
Genomic Imprinting Disorders
Genomic imprinting is where specific genes are expressed in a parent-of-origin manner via DNA methylation of either the maternal or paternal allele. Normally, we get two working copies (i.e., alleles) of each gene – one from each parent. However, in imprinted genes, you only get one working copy, either from your mother or father. For example, H19 is expressed only from the mother’s allele and is “shut off” on the father’s side (i.e., paternally imprinted). Genomic imprinting also involves the methylation of transposable elements (i.e., jumping genes) to maintain the integrity of the genome during gametogenesis. These epigenetic marks are established in the germ line and are generally maintained through mitotic divisions. However, sometimes these imprint genes are not properly methylated and lead to severe genetic diseases including Prader-Willi syndrome, Beckwith-Wiedermann syndrome, Silver-Russell syndrome, and Angelman syndrome. Although we’re still learning the full extent of the impact of these imprint genes, it’s very clear that they are critical for early brain development. In addition to causing neurological problems in children, improper imprinting may impact very early development (i.e., embryogenesis) and make it difficult to conceive (see below).
Male Factor Infertility (MFI)
Several studies have looked at specific genomic imprint loci to determine if aberrant DNA methylation at those locations is associated with MFI. Considering the importance of imprint acquisition during spermatogenesis, Marques et al. studied specific imprinted genes in different sperm populations. They compared the paternally methylated H19 differentially methylated region (DMR) and unmethylated MEST DMR in spermatozoa from fertile and infertile men. A loss of methylation in H19 was found in men with poor semen parameters, and an association between H19 methylation decrease and decreased sperm count was observed. An unexpected and abnormal methylation state of MEST DMR was also observed in oligozoospermic patient sperm. Using bisulfite conversion coupled with pyrosequencing, Boissonnas et al. performed a quantitative analysis of methylation levels at each CpG position included in IGF2 and H19 DMRs (47 different CpGs) in a population of normozoospermic and oligozoospermic men. They confirmed that the drastic loss of methylation restricted to IGF2 DMR2 (primary IGF2DMR) and H19 DMR correlated with the severity of the oligozoospermia. These data may suggest that some DMRs are more sensitive to methylation defects than others.
Poor Early Development (Embryogenesis)
In addition to problems with semen parameters, aberrant DNA methylation in sperm has been associated with poor IVF outcome, suggesting that sperm methylation marks affect early embryogenesis. Research by El Hajj et al. suggests that improper methylation of repetitive elements may be linked to recurrent pregnancy loss. Additionally, Benchaib et al. reported that among IVF patients, global sperm DNA hypomethylation is correlated with poor pregnancy outcomes. These data reveal an epigenetic landscape that appears programmed for use in the embryo with hypomethylated DNA at developmental promoters. Taken together, these data suggest that DNA methylation may play an essential role in both sperm function and early embryo development.
The current literature provides strong evidence for the importance of appropriate DNA methylation for both male fertility and IVF outcome. This essential epigenetic mark has tremendous utility in the maintenance of gene activation and repression. Inappropriate methylation clearly results in abnormal phenotypes. Developing diagnostic technologies that recognize these marks could greatly improve IVF outcome and well as accurately diagnosing men’s fertility status.